INTRODUCTIONStudies that evaluate the effects of hearing privation and stimulation in the inhibitory circuits (CHANG et al., 2003; KAPFER, 2002; KANDLER, 2004; RAJAN, 2002; SALVI et al., 2000; VALE et al., 2003) and in hearing perception (BOÉCHAT, 2004) have showed that the development and working of hearing system are associated to the quality and quantity of hearing input.
It is quite clear the relation between cochlear lesion, tinnitus development and intolerance to sounds. From 78% to 90% of patients with tinnitus presents alterations on audiometric thresholds (FUKUDA, 1990; MARTINS, 1991; COELHO et al., 2004). About 66% of patients with tinnitus have some grade of intolerance to sounds, and 33% from those searches specific treatment to hyperacusis, and 86% of patients with hyperacusis has tinnitus (JASTREBOFF and JASTREBOFF, 2004). Among patients searching treatment to hyperacusis, 53% also presents some grade of hearing loss (JASTREBOFF and JASTREBOFF, 2004).
The target of this study is to revise late findings about the effects of hearing privation on inhibitory circuits of central auditory nervous system and relate these findings with studies about tinnitus and hyperacusis with clinical, otorhinolaringology and phonoaudiology practice.
LITERATURE REVISIONIn order to help readers to understand the revised studies about the effects of hearing privation on inhibitory circuits of central auditory nervous system, we will briefly revise the basic concepts of the central auditory nervous system functioning (ROMAND, 1992; ROUILLER, 1992 and FERES, 1998).
The study of separate fiber physiology of central auditory nervous system shows that each fiber responds to a narrow and defined band of frequency. The analysis of frequency to which only one fiber responds and the thresholds of each response are called fiber tuning curve. In this curve, the most sensitive point of fiber, i.e., in which despolarization occurs with smaller threshold, is called characteristical frequency of fiber. From stimulation threshold of nervous fiber, the number of potentials of action grows according to the increase of stimulus intensity, reaching the saturation point, which is the maximum one. After that, the saturation point and the enlargement on stimulus intensity do not increase the frequency of potentials of action. The response band which goes from the threshold to the saturation point is called dynamic domain of fiber. The characteristical frequency, the spontaneous activity and the dynamic domain are features that provide an identity to each fiber of hearing nerve (ROUILLER, 1992).
In each nucleus of central hearing pathway, the properties of hearing neurons responses change into an afferent sense, showing that the signal is processed in a sequential and progressive manner. There are also efferent projections in almost all points of hearing pathway, what suggests that the nuclei are not independent; on the contrary, they are under control of superior centers in the pathway. In other words, the processing of hearing information is possible due to the cooperation of different parts of hearing system (ROUILLER, 1992).
The electrical signs that carry acoustic information are transmitted among neurons through synapsis. In the hearing pathway, most of synapsis is conducted by neurotransmitters (chemical synapsis), what makes the sign change all along its way, according to the necessity, if there are plastic changes in the system. Synapsis conducted by glutamate neurotrans¬mitter, for instance, are called exciting synapsis, because they intensify the possibility for postsynaptic neurons generates a potential of action, while the ones conducted by GABA and glycine, for instance (called s GABAergic and glycinergic), are considered inhibitory.
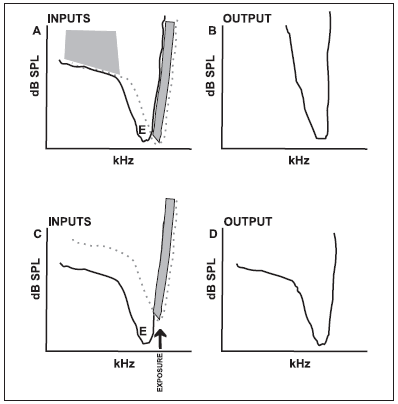
In the hearing, visual and somatosensory systems, the mechanisms of afferent inhibition start from the same area that provokes excitement on neurons (RAJAN, 2001). The inhibition occurs when quantity of inhibitory information is larger than the exciting one in the reception field. Thus, the points where inhibition surpasses areas of excitement of tuning curve of hearing fiber, there will be a masking (false impression) of these exciting responses to specific frequencies (Pictures 1A and 1B).
The exciting circuits compose 60% of the central nervous system. Therefore, the mechanisms that surround plasticity and development of inhibitory circuits of the central nervous system is still little understood (CHANG et al., 2003), though they are composed by a complex neuronal, morphological and functional chain. In the hearing pathway, however, the inhibitory circuits are organized and precise, and work as a model for the study of inhibitory function (KANDLER, 2004). Opposed to what occurs in topographic maps from other sensorial system, which has exciting synapsis (conducted by glutamate and acetylcholine), the organization of tonotopic map of hearing system has many clearly inhibitory pathways, conducted by glycine, GABA or both.
Picture 1. Schematic illustration of inhibition model by section. The pictures from above are from normal ear. The pictures from below are from ear that suffered acoustic trauma. A: tuning curve showing the interaction between exciting (solid line) and inhibitory (doted line) inputs of a normal neuron. Characteristical frequency of inhibitory input is placed in a frequency little above of characteristical frequency of exciting input. A normal hypothetical neuron produces an exciting response (open area above exciting tuning curve) if exciting input has lower threshold than the one from the inhibitory input. Responses are inhibited or masked (shadowed area), if inhibitory input has the same or inferior threshold as the one from the exciting input. B: The hypothetical exciting output of normal neuron in A. C: The same model is displayed, but threshold from inhibitory input is high as a result of an acoustic exposure at a tone that traumatizes the frequency above exciting characteristical frequency and close to inhibitory characteristical frequency. D: The same as B, but with a neuron output changed by the traumatizing tone. It is important to notice that the output of exciting responses is expanded, especially in the curve areas that respond to low frequencies. (Picture by SALVI et al., 2000, with author permission).
The most studied areas of hearing pathway that have inhibitory function are: medial superior olive (MSO) (which has an important role on sonorous localization); medial nucleus of the trapezoid body (MNTB), and lateral superior olive (LSO) (MNTB sends clearly inhibitory information from contralateral ear to LSO, which interacts it with exciting information of ipsilateral ear) and inferior colliculus (the largest center of subcortical hearing integration).
Late researches suggest that inhibitory circuits perform an important role in intensity perception, in hearing distinguishion, in tinnitus and in hyperacusis.
Physiological evidences of central auditory nervous system plasticity by hearing privation
Plasticity is a biodynamic phenomenon which causes physiological, biochemical and/or anatomical changes of the central neurons due to the demand of acoustic information transmission. Thus, the hearing system organizes itself when there is a variation on hearing input, by either input reduction, in cases of cochlear functions or by increasing, when there is another hearing input (during postnatal development and after cochlear implantation or hearing aid, for instance).
The development and maintenance of fibers and other structures of central auditory nervous system depend on the interactions between pre and postsynaptic neurons. KAPFER et al. (2002) studied the development of the first structure of the central auditory nervous system that is responsible for the sonorous localization of low frequencies in horizontal plan, the medial superior olive (MSO) in guinea pigs. MSO receives exciting (glutamate) and inhibitory stimuli (glycine) from the two ears and compares the difference in interaural time. As this difference can be only some microseconds, glycinergic synapsis needs to be highly precise. In order to optimize postsynapitc temporal precision, the glycinergic inputs occur close to neuron discharge zone (cellular bloody or proximal dendrite). Nevertheless, this subtility is only achieved soon after two weeks of life with hearing stimulation. In newborn guinea pigs, in young ones with unilateral cochlear ablation and in the ones who received unidirectional sonorous stimulus, it was observed that glycinergic synapses are regularly distributed in the cellular body, proximal and distal dendrites. Therefore, the development and organization of inhibitory neuronal chain, which is fundamental for refined tuning, is dependent on the activity (KAPFER et al., 2002).
When there is loss of pre synaptic elements, the inter and intracellular events change. In the post-synaptic neurons, it can occur alterations of their structure, function and metabolism, what it might lead to their death (ZIRPEL et al., 1997). In this way, cochlear lesion can cause severe changes in exciting responses on cortical neurons that supply the hearing system.
When there is a partial lesion of the cochlea, the central areas which were responsible for injured cochlear areas are now occupied by inputs from preserved areas of the cochlea. In other words, tonotopic maps from the whole central auditory system can be changed by the expansion of frequencies representations whose perception threshold is normal or better (WILLOT, 1996; RAJAN, 2001).
Another effect on the central auditory nervous system after cochlear lesion is the unmasking process, i.e., the manifestation of exciting inputs that used to be inhibitory (RAJAN, 2001). In this way, a small lesion, either permanent or temporary on the receiver organ, or such condition that simulates hearing loss, can lead to a loss of immediate inhibition around only one cell of the central auditory nervous system (Pictures 1C and 1D) (RAJAN, 2001; SALVI et al., 2000).
WANG et al. (1996) evaluated the responses of the nucleus of inferior colliculus and determined its frequency curves (tuning curve). They also observed fibers which were excited only by a narrow band of frequency at 8 kHz, and the discharges of this frequency increased up to 20 dB above threshold and decreased to higher intensities. After the contralateral ear being exposed at 16 kHz (one eighth above) at 107 dB NPS per 20 minutes the threshold of characteristical frequency of the neuron and the response for high frequencies did not change. However, the threshold to low frequencies increased up to 25 dB in frequencies of 4 to 5 kHz and the fiber started to respond to lower frequencies (500 Hz). Such changes occurred in about 40% on neurons of inferior colliculus, i.e., the largest part of the fiber of inferior colliculus was affected by traumatic exposure. Affected and non-affected cells can be distinguished by their functional features, what suggests the latter does not have important lateral inhibitory input.
In a similar study, SALVI et al. (1996) evaluated the consequences of exposure to intense sounds on dorsal cochlear nucleus. Findings showed that neurons with narrow exciting areas (that respond to few frequencies) and inhibitory areas had the same functional changes found on inferior colliculus, while cells with ample responses to frequencies had small changes.
The effects of lesion of internal ciliated cells by carboplatin over electric activity on round window, on inferior colliculus and on primary hearing cortex were researched through auditory evoked potential in these three areas (WANG et al., 1997). The data show a reduction of 50% of the auditory evoked potentials of round window, little or none alteration on input and output function of inferior colliculus for weak and moderate intensities and reduction reaching up to 50% for stronger intensities, and enlarging the potential amplitude of hearing cortex, with large inter-animal variability. The top of amplitude of auditory evoked potentials on hearing cortex (above normal condition) was reached some weeks after lesion and then reduced a little. Findings suggest that there is a gain control between cochlea and brainstem that compensates or reduces the effects of one lesion of internal ciliated cells. There seems to be a gain control of hearing cortex that aims to adjust hearing gain in order to keep it in normal operational band (WANG et al., 1997; SALVI et al., 2000).
Clinical evidences of central auditory nervous system plasticity by hearing privation and stimulation
With the purpose of comparing the development on intensity perception between users and non-users of devices of individual sonorous amplification, PHILLIBERT et al. (2002) evaluated 18 subjects with symmetrical bilateral sensorineural hearing loss (SNHL) (aging from 64 to 90 years), 9 non-users and 9 users of devices of individual sonorous amplification (for a period from 1 to 5 years) on both ears (digital and with gain control), paired by age, sex and loss grade. Authors applied loudness scale and Discrimination-limen for intensity, which evaluate the ability of the individual in detecting little intensity differences. Results showed expressive differences on both tests. Users of individual sonorous amplification can notice smaller differences of intensity and classify as "ok" the sounds which non-users classify as "strong". Thus, there seems to be changes on intensity codification of the subjects who are users of devices of individual sonorous amplification.
Yet the sensation of tinnitus intensity does not seem to be changed due to time of hearing loss settling. OCHI et al. (2003) did loudness and tinnitus pitch measuring in patients with unilateral hearing impairment and unilateral tinnitus (N=132). The subjects were divided according to period of time of the symptoms. One of the groups was formed by patients with tinnitus and acute hearing impairment (sudden deafness less than 7 years) and the other one was formed by patient with chronic tinnitus and chronic hearing impairment (for more than 3 months and of idiopathic origin). Authors related loudness and tinnitus pitch measurement to hearing loss settling. The differences statistically significant between the two groups were: age average, lower to the group of sudden deafness and sudden tinnitus (average of 44.0 years ± 13.6, against 54.0 years ± 15.9 in the group of chronic hearing impairment and chronic tinnitus) and tinnitus pitch distribution. Tinnitus pitch distribution from the group of sudden deafness and sudden tinnitus was diffuse, with average of 5.81 kHz (SD = 3.22 kHz), in frequencies a little lower from those in which the largest difference of audiometric threshold between the affected and non-affected ear lay. In the group of chronic tinnitus and chronic hearing impairment, tinnitus pitch was higher, with an average of 2.96 kHZ (SD = 3.30 kHz) and with two peaks of tinnitus pitch distribution, one was little below frequency with greater difference of audiometric threshold between affected and non-affected ear and other was 1/3 of eighth above tinnitus pitch. Therefore, it was observed that tinnitus pitch corresponded to 0.2 eighth below frequency of greater hearing asymmetry between two ears, but its relation with time of hearing loss settling is different, what suggests that hearing cortex of patients with chronic hearing impairment and chronic tinnitus suffered some reorganization.
BOÉCHAT (2003) examined the interference of privation and stimulation time on variation of hearing sensitiveness to clean tones and percentual rate of speech recognition. 72 subjects (aging from 10 to 86 years) were examined with bilateral or unilateral asymmetrical SNHL. 43 from those were users of devices of individual sonorous amplification and 29 were non-users in the worst ear. Considering three audilogic evaluations (initial, intermediate and final), it was analyzed the variations to clean tone of sensitiveness and percentual rate of speech recognition between groups in 6 years at most in relation to stimulation and privation time, asymmetry grade between ears and loss grade. As the group of users of devices of individual sonorous amplification had smaller variation of tonal thresholds and better percentual rate of speech recognition, the author concluded that it occurred both plasticity by hearing privation with non-users of devices of individual sonorous amplification in the worst ear and secondary plasticity after hearing stimulation with users of devices of individual sonorous amplification. It also was observed that in the first years after introduction of hearing stimulus there is a gain when using devices of individual sonorous amplification, while the negative effects of privation gradually present itself in homogenous manner along time. Boéchat considered that stimulus quantity and quality determine the development and maintenance of central auditory system and pointed out the importance of binaural hearing stimulation.
Silence effects on normal hearing pathway
Tinnitus is usually noticed by patients when they are in quiet places without doing anything that may call their attention. The most common example of this situation is when the individual is in bed, though the discomfort with tinnitus perception can make their sleep difficult or even prevent it.
The studies which evaluated the silence effect in the hearing pathway in individuals with normal hearing are rare. The study of HELLER AND BERGMAN (1953) is still remembered for having demonstrated that 94% of the subjects without complaint of hearing loss (with no audiometric proof) presented tinnitus after being in a quiet booth for five minutes. In the same type of study, TUCKER et al. (2005) evaluated the silence effect when appearing of tinnitus perception in 120 adults (60 men and 60 women) aging from 18 to 30 years with normal hearing. After being in an acoustically designed booth for 20 minutes, the individuals were questioned on any sounds they might have heard during quiet time. 64% of the subjects (n=77) reported having noticed some sounds in quiet time (including heartbeats), with expressive difference between races, and not gender (78% of Caucasians and 38% of black noticed the tinnitus).
If we consider that most of people remain in relatively quiet places, at least in sleeping time, we could expect more reports on tinnitus perception in normal-hearing individuals than usual. Perhaps the information provided to the participants might have called their hearing attention though, even not having the knowledge that this sound could come from them, they were previously informed they could hear some sounds when in the booth.
Current studies suggest that patients disturbed by tinnitus should receive, besides previous disease treatment, competitive sounds that reduce the contrast between tinnitus and the environment (JASTREBOFF and HAZELL, 1993; HAZELL, 2002; SANCHEZ, 2002; SANCHEZ and FERRARI, 2003; KNOBEL et al, 2004; JASTREBOFF and JASTREBOFF, 2004).
Tinnitus Retraining Therapy was developed in early 90´s with the purpose of encouraging the habit of induced reactions by tinnitus and by intolerance to sounds (JASTREBOFF and HAZELL, 1993). It consists of individual sections of sound orientation, follow-up and improvement. The type of sound improvement is recommended according to hearing complaints from the patient (grade of tinnitus discomfort, type of intolerance to sounds and grade of hearing loss, if exist). In general terms, it is recommended the use of light, continuous and monotonous sounds to all kinds of patient. These sounds can be present in the environment (music, sounds of nature, water fountains, fans, etc), directly into acoustic meatus or by sonorous amplification of environmental sounds through devices of individual sonorous amplification, to people with hearing impairment. In Tinnitus Retraining Therapy, the adaptation of sound generator devices or devices of individual sonorous amplification (if there is bilateral hearing impairment) should always be bilateral, even if tinnitus of patient is unilateral. Staying in quiet places as well as using hearing protectors in places with no intense noise, are not recommended (JASTREBOFF and HAZELL, 1993; JASTREBOFF and JASTREBOFF, 2004).
DISCUSSIONIn physiological state, tinnitus can be explained by any changes in neural synchrony or in standard of electrical discharge of spontaneous neural activity of hearing pathways (EGGERMONT, 2003). The acute appearing of tinnitus after cochlear lesion can be explained by loss of lateral inhibition, that would unmask exciting responses of neural circuits previously inhibited (SALVI, 2000; RAJAN, 2001). Thus, unmasking new exciting responses suggests that inhibition loss has a critical permissive role (RAJAN, 2001; OCHI et al, 2003). Yet, chronic tinnitus and intolerance to sounds can be related to a gradual hyperactivity of hearing cortex as a consequence of cochlear lesion (SALVI, 2000) and cortical reorganization (OCHI et al, 2003).
The presented studies showed plastic changes in portions of the hearing pathway that codify intensity. Such changes occur due to quantity and quality of acoustic stimulation and reflect in intensity perception. Findings of SALVI (2000) suggest that there is a gain control between cochlea and brainstem that compensates or reduces the effects of lesion of internal ciliated cells. There seems to be a gain control of hearing cortex that aims to adjust hearing gain in order to keep it in normal operational band. It is likely that in patients with hyperacusis, this gain control is not working properly, what could cause abnormal perception of light and moderate sounds (JASTREBOFF and JASTREBOFF, 2004).
JASTREBOFF and JASTRBOFF (2004) purpose that hearing privation by reduction of cochlear output can either cause tinnitus or strengthen a pre-existent one. This relation seems to be associated to plastic changes that occur in central auditory system after lesion of receiver organ, and to the direct consequences of cochlear lesion (SALVI, 2000).
The findings of BOÉCHAT (2003) give special attention to precocious aids and to balanced stimulation between ears and even suggest that symmetric bilateral hearing losses with adaptation of monoaural device of individual sonorous amplification can change into asymmetrical one due to asymmetrical stimulation. This information also supports recommendation to Tinnitus Retraining Therapy of which the use of sound generators is always bilateral and symmetry between ears is kept at its maximum (JASTREBOFF and HAZELL, 1993; JASTREBOFF and JASTREBOFF, 2004). In our clinical practice we observe that patients with tinnitus who adjust devices of individual sonorous amplification or sound generator only in the ear affected by tinnitus (unilateral tinnitus) or in the ear with the worst tinnitus (bilateral tinnitus) start to notice tinnitus in contra-lateral ear. We believe that, in short term, tinnitus either improvement or masking in the worst ear make tinnitus perception easy, which was subclinical in the other ear.
The deep search of the origins which rules the functional organization of inhibitory pathways of hearing system can provide the understanding of abnormalities in the organization of inhibitory circuits that seem to be present on tinnitus and on intolerance to sounds. From that, new therapeutical strategies can be developed.
FINAL COMMENTSThe literature has been providing evidences of plasticity of hearing pathways which suggest that otorhinolaryngology and phonoaudiology everyday practice should be revised in relation to situations little considered. Thus, we point out the benefits of precocious diagnosis of situations of hearing privation, tinnitus, intolerance to sounds and also to its treatment, even if it is by bilateral precocious hearing aid in cases of hearing loss, or to the use of bilateral sonorous stimulation and symmetric in cases of tinnitus and intolerance to sounds.
BIBLIOGRAPHY1. Boéchat EM. Plasticidade do sistema auditivo quanto à sensibilidade auditiva para tons puros e respostas para a fala na deficiência auditiva neurossensorial. São Paulo, 2003,175p. (Tese de Doutorado - Faculdade de Medicina da USP).
2. Chang EH, Kotak VC, Sanes DH. Long-term depression of synaptic inhibition is expressed postsynaptically in the developing auditory system. J Neurophysiol, 2003, 90(3):1479-88.
3. Dauman R, Bouscau-Faure, F. Assessment and amelioration of hyperacusis in tinnitus patients. Acta Oto-Laryngologica, 2005, 125: 503-509.
4. Féres MCLC. Estudo das respostas plásticas de núcleos cocleares secundárias à deafarentação unilateral por ablação de cóclea em ratos Wistar neo-natos. [tese - doutorado - Universidade Estadual de São Paulo]. Ribeirão Preto, 1998.
5. Hazell JWP, Sheldrake JB, Graham RL. Decreased sound tolerance: predisposing factors, triggers and outcomes after TRT. In: International Tinnitus Seminar, 7. 2002. Fremantle, Australia. Proceedings of VII International Tinnitus Seminar, Perth, Australia: The University of Western Australia, 2002. p.255-61.
6. Jastreboff P, Jastreboff MM. Tinnitus Retraining Therapy (TRT) as a Method for Treatment of Tinnitus and Hypercausis Patients. J. Am. Acad. Audiol., 2000, v.11, n.3, p. 162-177.
7. Jastreboff P, Jastreboff MM. Decreased Sound Tolerance. In: SNOW J. Tinnitus: Theory and Management. Hamilton: BC Decker, 2004. p. 8-15.
8. Kandler K. Activity-dependent organization of inhibitory circuits: lessons from the auditory system. Curr Opin Neurobiol, 2004, 14: 96-104.
9. Kapfer C, Seidl AH, Schweizer H, Grothe B. Experience-dependent refinement of inhibitory inputs to auditory coincidence-detector neurons. Nat Neurosci, 2002, 5:247-253.
10. Ochi K, Ohasi T, Kenmochi M. Hearing Impairment and tinnitus pitch in patients with unilateral tinnitus: comparison of sudden hearing loss and chronic tinnitus. Laryngoscope, 2003, 113: 427-431.
11. Phillibert B, Collet L, Vesson J-F, Veuillet E. Intensity-related performances are modified by long-term hearing aid use: a functional plasticity? Hear Research, 2002, 165: 142-151.
12. Rajan R. Plasticity and inhibition in the receptive field of primary auditory cortical neurons after limited receptor organ damage. Cerebral Cortex, 2001, 11:171-182.
13. Romand R. Aspects anatomo-fonctionnels du noyau cochléaire. In: Romand R. (Ed). Le systeme auditif central. Anatomie et physiologie. Les Éditions Inserm, Paris, 1992, 51-108.
14. Rouiller EM. Organisation fonctionnelle des voies auditives. In : Romand R. Le Systeme Auditif Central. Les Éditions Inserm, Paris, 1992, 1-49.
15. Salvi RJ, Wang J, Ding D. Auditory plasticity and hyperactivity following cochlear damage. Hear Research, 2000, 147: 261-274.
16. Tucker DA, Phillips SL, Ruth RA, Clayton WA, Royster E, Todd AD. The effect of silence on tinnitus perception. Otolaryngol Head Neck Surg, 2005, 132(1):20-4.
17. Vale C, Schoorlemmer J, Sanes DH. Deafness disrupts chloride transporter function and inhibitory synaptic transmission. J Neurosci, 2003, 23:7516-7524.
18. Wang J, Salvi RJ, Powers N, Plasticity of response properties of inferior colliculus neurons following acute cochlear damage. J Neurophysiol, 1996, 75: 171-183.
19. Wang J, Powers NL, Hofstetter P, Trautwein P, Ding D, Salvi R. E¡ects of selective inner hair cell loss on auditory nerve fiber threshold, tuning and spontaneous and driven discharge rate. Hear. Res., 1997, 107:67-82.
20. Willot JF. Physiological plasticity in the auditory system and its possible relevance to hearing aid use, deprivation and acclimatization. Ear Hear, 1996, 17:66-77.
21. Zirpel L, Lachica EA, Rubel EW. Afferent regulations of neurons in the avian cochlear nucleus: intracellular mechanisms and signal transduction pathways. In: Berlin C. (ed). Neurotransmission and hearing loss - Basic science, diagnosos and management. San Diego: Singular Publing Group, 1997, p. 77-89.